Passiflora ambigua
P. ambigua is a forest species, growing in the canopy, treefall clearings and along trails. The flowers and fruits are large and showy, with fruits 5-7 cm diameter and 12-15 cm long. The simple, drip-tipped leaves look like many other leaves in the forest understory, but the reaching shoot tips with extended tendrils are fairly easy to see. The petiolar nectaries are large and attractive to Ectatomma and other ants. Stems, both small and large (up to 5cm) are round and woody. Range: Southern Mexico to Peru and Venezuela. Wet forest 0 to 1400 meters elevation; in primary forest and forest edges. Another similar species, P. nitida (range: Costa Rica to Brazil) , is also recorded from La Selva from sites in the upland forest. Its leaves are distinguished from P. ambigua in that the nectar glands are located at the apex of the petiole, not the middle as with P. ambigua. Member of section Laurifoliae, subgenus Passiflora.
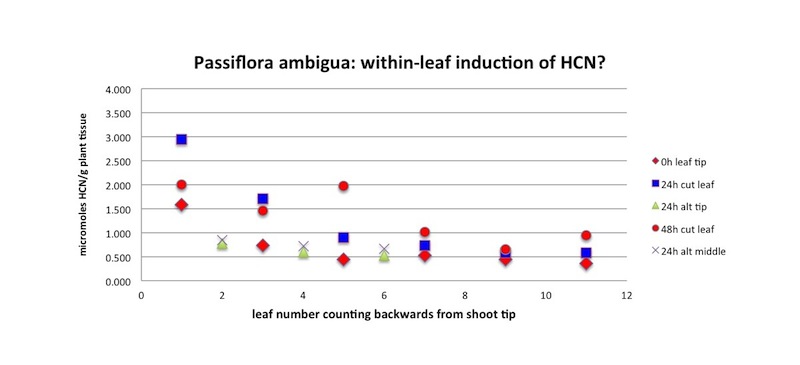
This species is cyanogenic, exhibiting a wide range of concentrations (0.03 - 10 microMole HCN/g plant tissue). Spencer (1988) reported that P. nitida and P. laurifolia, close relatives of P. ambigua, contain the simple monoglycoside sulfated cyclopentyl cyanogens Tetraphyllin B sulfate, Epitetraphyllin B sulfate, and Passilaurifolin. It also contains substantial amounts of alkaloids (Smiley and Wisdom 1985). As can be seen in the graph below, P. ambigua possesses a "fast reaction" enzyme/substrate system for producing HCN gas when the leaves are crushed. However, there is some evidence of an additional slow-release process in the young leaves only.
P. ambigua is one of the three most variable Passiflora species in terms of cyanogenesis. Like P. auriculata and P. biflora, HCN production in new leaves ranges over two full orders of magnitude (100x; see Figure 5b). Analysis of HCN production in over 100 leaves belonging to over 30 branches from 16 different plants revealed that about 1/3 of the variation may be attributed to differences between plants, another 1/3 to differences among branches on the same plant, and about 1/3 to differences between leaves on the same branch (Figure 5d)
The group-feeding, obligately monophagous Heliconius doris feeds on P. ambigua. As a feeding specialist it may possess the ability to sequester and metabolize the HCN-producing glycosides in the host plant. However, P. ambigua glycosides are probably sulfated and may have different properties than the simple cyclopentenoid cyanogens that have previously been shown to be sequestered by other Heliconius (see Table 5). Multiple H. doris females oviposit "rafts" of small yellow eggs, as many as 800 on one leaf.
It is notable that H. doris females frequently lay eggs on the large P. ambigua growing in the garden next to the laboratory. This plant is not highly cyanogenic, averaging about 0.1 μM/g HCN, yet seems to be a preferred host. Perhaps, contrary to prediction, H. doris does not seek out plants rich in cyanogenic glycosides.
Heliconius cydno lays her eggs on the tendrils and leaf tips of this plant. The large petiolar nectaries are highly attractive to ants such as Ectatomma sp. If the plant is frequented by Ectatomma ants (i.e. near an Ectatomma nest), the probabiity of caterpillar survival is very low. If Ectatomma is rare or absent from the plant, then many caterpillars will survive. Two other generalist Heliconius, H. hecale and H. ismenius, also oviposit on the tendrils and leaf tips.
P. ambigua is often fed upon by the generalist Blue Flea Beetle (Monoacra violacea) and Black-tibia Flea Beetle (Parchicola "black tibia"). Yellow-tibia Parchicola has also been found on P. ambigua, but Parchicola sp. eggs and larvae have not been found. On P. oerstedii, Yellow-tibia Parchicola eggs are laid on the undersides of old leaves near ground level, and larvae feed on the basal stems and perhaps surface roots.
The Yellow Ptocadica Flea Beetle (Ptocadica sp. "yellow") feeds on P. ambigua, although it may also be found on P. oerstedii, and less commonly, P. vitifolia. Feeding trials indicate that Yellow Ptocadica adults may feed successfully on several species of Passiflora such as P. biflora and P. auriculata. Eggs and larvae are, as yet, unknown, but like other Ptocadica are probably found on the under sides of the leaves.










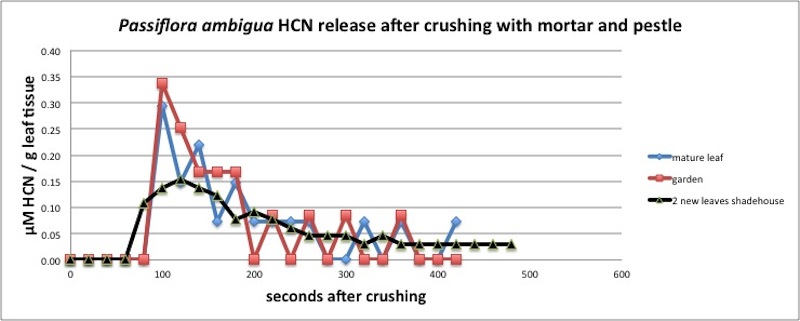

Clearly a "fast reaction" completed within 5 minutes, regardless of the actual amount of HCN.